RESEARCH

OVERVIEW
The Carter lab studies evolutionary biology, animal behavior, the economics of public health, and whatever else attracts Dr. Carter's attention.
Students in the lab use
comparative (i.e., analyzing published data sets),
theoretical (i.e., math and computer simulations) and
empirical (i.e., lab experiments, mainly with
Drosophila) approaches to address these topics.
The broad
evolutionary question we study is what influences the evolvability, the capacity to evolve, of organisms and species. Evolvability can be influenced by characteristics of populations (e.g., population size) or individuals (e.g., developmental stability) and we study both.
The lab also studies the
behavior of
Drosophila. Recent studies have mainly focused on the mate choice behaviors of both female and male individuals.
The lab has also examined the
economic relationship between governmental research spending on various medical conditions and their burden or cost to society as a whole.
Other studies that students have worked on include the effects of different fonts on learning and examining published data sets for evidence of scientific fraud.
EXAMPLES OF PAST AND CURRENT PROJECTS
Drosophila Mating Behavior
Drosophila melanogaster is a model organism used in evolutionary biology and genetics as well as in many biomedical studies of behavior and neurobiology. Understanding the factors that influence the behaviors of members of this species is therefore essential for understanding this very important organism. The mate choice behaviors of
Drosophila are a subject of study in my lab and we have two publications on this topic so far; additional projects are in progress and/or ready to begin.
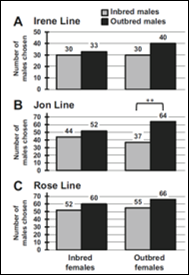
Working with two students I investigated the influence of inbreeding on the mate choice preferences of females in two species of
Drosophila. The degree to which inbreeding altered both the preferences of inbred or outbred females when selecting inbred or outbred males was measured and our evidence suggested that outbred females were more selective, contrary to predictions arising from previous published studies. The student co-authors on
that publication were each accepted into a Ph.D. program in 2014 in part due to their experiences with this project.
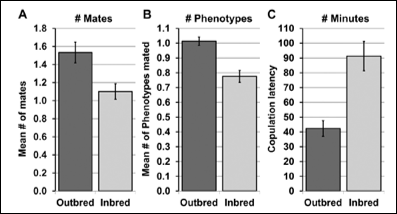
Working with a graduate student and a number of undergraduates I investigated the influence of inbreeding on the mate choice preferences of females in two species of
Drosophila. The degree to which inbreeding altered promiscuity of inbred vs outbred females was measured and our evidence suggested that inbred females were less promiscuous, contrary to predictions arising from previous published studies. The student co-author on
that publication was accepted into a Ph.D. program in 2015 in part due to her experience with this project.
Morphological Asymmetry
Most organisms are bilaterally symmetric and deviations in observed symmetry therefore arise from errors in development. The degree to which the development of an organism is disrupted by stressful factors in the environment or resistant to these perturbations can be estimated from measuring the non-directional symmetry, the "fluctuating asymmetry" (FA), observed. Selection on symmetry can be estimated from comparing young-of-the-year to the adults that survive. The degree to which environmental quality or stress can influence development can be estimated from comparing the FA in regions that differ in their environmental conditions. In 2016 I published
a paper with a graduate student and a fellow CSULB faculty member comparing FA for young and old fish in two regions that differed in pollution levels.
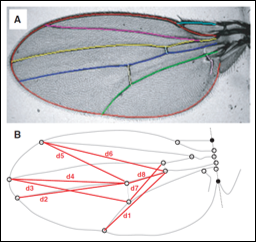
Artificial selection is a powerful technique that allows details of the genotype phenotype map to be studied. The lab has the facilities to raise and measure wing traits in large numbers of
Drosophila while they are still alive, allowing artificial selection on these traits to be performed very efficiently. Two studies with published results are detailed below; additional projects are in progress and/or ready to begin.
Fluctuating asymmetry is caused by slight errors and imperfections during development leading to morphological traits differing between the two sides of an organism. The presence or absence of genetic variation that may predispose individuals to higher or lower levels of such developmental errors (arising from differing degrees of developmental stability) is potentially relevant for understanding developmental disorders in humans such as cleft palate or club foot as well as being inherently interesting as a subject of natural selection. The NIH funded a study in which I worked with a team of undergraduate students to perform an artificial selection experiment on the fluctuating asymmetry of wing vein traits, the first of its kind.
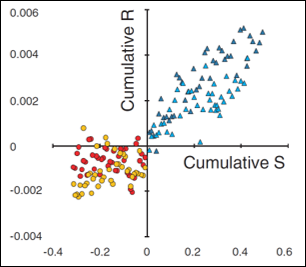
Our study showed responses to both increase and decrease fluctuating asymmetry; demonstrating the existence of genetic variation for developmental stability directly for the first time.
Directional asymmetry exists when the mean value of one side differs from that of the other side (i.e., average right is larger than average left) and can be seen to evolve in nature, but previous studies using artificial selection reported a lack of selectable genetic variation in laboratory studies. I worked with an undergraduate student to perform an artificial selection experiment on the directional asymmetry of wing vein traits that was designed to lack many of the methodological problems present in previous studies.
Our study also showed no response, contributing to the body of knowledge regarding this mysterious trait. The student co-author on that project was accepted into a Ph.D. program in 2008 in part due to her experience with this project.
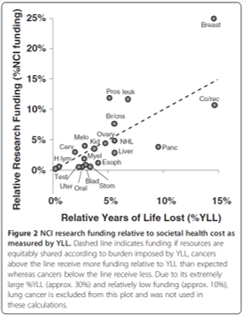
Statistical analyses of the economics of cancer research
Cancer kills many individuals and in order to address this peril, governments fund grants and institutions specifically devoted to cancer treatment research. It is in the best interest of government agencies to ensure this money is spent effectively and efficiently; since financial resources are limited these resources should therefore be devoted to cases in which the greatest benefit is likely to be realized. For example, the NIH recommends that research effort be compared to societal burden when making funding decisions, but studies explicitly comparing cancer research funding to societal burden are few in number and often group all types of cancer together or include other diseases with quite different etiologies which obscure comparisons. Quantitatively comparing the burdens arising from various cancers to research funding devoted to them therefore has the potential to greatly improve public health by optimizing funding allocation. The use of publicly available data allows comparative studies to be performed that have the potential to address this and I have two projects completed on this topic to date; additional projects are in progress and/or ready to begin.
In 2012 I published
a paper with a former student that analyzed US DALY and YLL values with regard to research funding and identified a number of cancers which appear to be relatively overfunded and underfunded. This information may prove to be of high value to government officials making funding allocation decisions in the future. The student co-author on that project was accepted into a M.S. program in public health primarily due to her experience with this project.
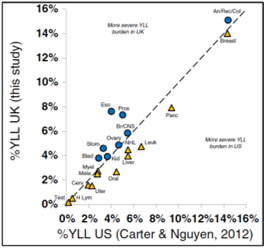
In 2015 I published
a paper with two former students comparing the funding allocation discrepancies in the UK to those we identified in the US and to previously reported UK values. The funding and burden patterns in the two countries were broadly similar.
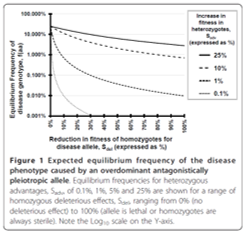
Antagonistic pleiotropy as an evolutionary mechanism for genetic disease
Many serious diseases have a genetic basis which, from a naive point of view, should have been selected against in the past; the sustained prevalence of a number of disease-associated alleles is therefore surprising. A more sophisticated evolutionary approach can yield information about what maintains genetic variation in situations in which we may expect it to be lost. I have completed one project on this topic to date; a follow up study using additional resources is available for interested students.
I worked with a student to survey the medical literature in order to determine whether sufficient evidence exists to argue that antagonistic pleiotropy (when a genetic variant has both positive and negative effects) may be responsible. We also used a population genetic model to examine the feasibility of antagonistic pleiotropy to act as a mechanism to maintain polymorphism for serious genetic disorders even if the benefits are subtle. We identified a number of examples of antagonistic pleiotropy where the deleterious effect, the beneficial effect, and the exact molecular cause have been demonstrated. We also identified putative cases in which there is circumstantial evidence or a strong reason to expect antagonistic pleiotropy in a genetic disorder. The population genetic model demonstrated that alleles with severe deleterious health effects can be maintained at medically relevant frequencies with only minor beneficial pleiotropic effects. If antagonistic pleiotropy is as common as our initial research suggests, this may explain why so many serious diseases, even apparently environmentally caused ones, have a genetic component. The student co-author on
that publication was accepted into medical school in 2013 in part due to his experience with this project.
Work on a follow-up project I conducted with two undergraduate student co-authors, where we identified many more examples, should be published soon.